Tổng hợp các kết quả nghiên cứu về FAdV (Fowl Adenovirus) ở Việt Nam
Bộ môn vi sinh vật – Truyền nhiễm
Trên thế giới cũng như các nước trong khu vực, FAdV đã được xác định lưu hành và là căn nguyên gây ra các tình trạng bệnh lý ở gà như: loét dạ dày tuyến (Lim & cs, 2012), tích nước xoang bao tim (hydropericardium) và viêm gan thể vùi (inclusion body hepatitis) (Choi & cs, 2012). Tuy nhiên, ở Việt Nam, theo hiểu biết của chúng tôi, kết quả trình bày dưới đây là nghiên cứu đầu tiên về sự lưu hành của FAdV ở đàn gà. Trên cơ sở đó, đã cung cấp cơ sở quan trọng cho các nghiên cứu tiếp theo về tình hình bệnh do FAdV gây ra.
Gà có kết quả dương tính FAdV biểu hiện một số bệnh tích như gan, lách xuất huyết, sưng to như minh họa ở hình 2.19 – 2.20.
 |
 |
Nghiên cứu được thực hiện tại Bộ môn Vi sinh vật- Truyền nhiễm, Khoa Thú y, Học viện Nông nghiệp Việt Nam. Để xác định sự lưu hành FAdV ở đàn gà nuôi tại Hà Nội và vùng phụ cận, đã sử dụng phản ứng PCR; sau đó tiến hành giải trình tự gen mã hóa protein hexon (gen hexon) của FAdV và phân tích một số đặc điểm dịch tễ học phân tử của FAdV.
Cặp mồi đặc hiệu dùng phát hiện FAdV được lựa chọn theo nghiên cứu đã công bố (Ganesh & cs, 2002), với trình tự được trình bày ở bảng 2.3
Bảng 2.3. Trình tự mồi đặc hiệu phát hiện và giải trình tự gen hexon của FAdV
| Mồi xuôi/ ngược | Trình tự 5’ – 3’ | Kích thước (bp) |
| FAVHL/ | GACATGGGGTCGACCTATTTCGACAT | 731 |
| /FAVHR | AGTGATGACGGGACATCAT |
Nhằm tìm hiểu đặc điểm dịch tễ học phân tử của FAdV serotype 4 (FAdV4) phát hiện được trong nghiên cứu này và trả lời câu hỏi: (1) nguồn gốc phát sinh từ đâu và (2) lưu hành ở Việt Nam từ khi nào, chúng tôi tiến hành phân tích đặc điểm dịch tễ học phân tử theo không gian (quốc gia). Cơ sở dữ liệu dùng cho phân tích là một phần trình tự gen mã hóa protein hexon các chủng FAdV4 (có trong ngân hàng gen, Genbank) tương đồng ít nhất 99% về trình tự nucleotide với chủng FAdV4- F5. Các bước thực hiện được tóm tắt như sau:
(1) Dùng công cụ BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) để tìm kiếm trình tự gen tương đồng 99% với đoạn gen so sánh,
(2) Sàng lọc và giữ lại các trình tự có đầy đủ thông tin về địa điểm phân lập (quốc gia) và năm phân lập,
(3) Dùng phần mềm BEAST để xây dựng lại quá trình phát tán của các chủng virus FAdV4 theo không gian và thời gian, với các tham số dựa theo kết quả nghiên cứu trước đây (Lemey & cs, 2009).
Thực hiện phản ứng PCR để xác định sự có mặt của FAdV trong các mẫu bệnh phẩm thu thập trong tổng số 125 mẫu bệnh phẩm của gà thuộc 14 trại có triệu chứng lâm sàng được kiểm tra thì tỷ lệ phát hiện được FAdV là 30 mẫu (tỷ lệ 20,69) (Lê Văn Trường & cs, 2020). Kết quả trên cho thấy trong các đàn gà nuôi tại khu vực Hà Nội và vùng phụ cận có sự lưu hành FAdV. Với đặc tính gây suy giảm miễn dịch thì sự lưu hành của FAdV trong đàn gà sẽ là một thách thức mới với người chăn nuôi tại Việt Nam.
2.1. Kết quả phân tích trình tự nucleotide gen hexon của FAdV
Trong 26 mẫu dương tính với FAdV, chúng tôi lựa chọn 14 mẫu để tiến hành giải trình tự gen, trong đó có 7 mẫu được giải trình tự thành công, trình tự của đoạn gen hexon của 7 chủng được trình bày ở hình 2.21.

Hình 2.21. Trình tự gen hexon của 07 chủng FAdV
Phân đoạn gen hexon từ nucleotide 361 đến 972. Dấu “.” biểu thị các vị trí có nucleotide giống nhau so với trình tự số 1 (chủng F5).
Kết quả ở hình 2.21 cho nhận xét chung: trình tự gen hexon của 7 chủng FAdV rất đa dạng. Các đặc điểm về trình tự gen của 7 chủng FAdV được tổng hợp như sau:
– 24/612 vị trí có đột biến mất nucleotide,
– 271/612 (chiếm 44,28%) vị trí không có thay đổi nucleotide (monomorphic),
– 317/612 (chiếm 51,80%) vị trí có thay đổi (đa hình nucleotide, polymorphic).
Trong các vị trí này, có 210 vị trí được xếp vào nhóm “vị trí có thông tin” (parsimony-informative site, là vị trí mà ở đó có ít nhất 2 sự thay đổi loại nucleotide và mỗi một loại thay đổi nucleotide xuất hiện ở ít nhất 2 trình tự khác nhau), bao gồm: 120 vị trí có 2 sự thay đổi nucleotide, 80 vị trí có 3 sự thay đổi nucleotide, và 10 vị trí có 4 sự thay đổi nucleotide. Với các trình bày nêu trên, có thể thấy rõ tính đa hình nucleotide của các chủng FAdV mà chúng tôi giải trình tự gen. Đặc điểm này là phù hợp với những hiểu biết về tính đa dạng genotype, serotype của FAdV đã được biết trên thế giới (Hess, 2000; Meulemans & cs, 2004).
Tính đa dạng về trình tự gen giữa các chủng FAdV còn được biểu diễn dưới dạng % tương đồng nucleotide (bảng 2.4).
Bảng Kết quả so sánh mức tương đồng nucleotide giữa 7 chủng FAdV
| F5 | F11 | F6 | F9 | F7 | F3 | F8 | |
| F5 | ID | ||||||
| F11 | 0,575 | ID | |||||
| F6 | 0,598 | 0,809 | ID | ||||
| F9 | 0,598 | 0,793 | 0,978 | ID | |||
| F7 | 0,609 | 0,875 | 0,817 | 0,811 | ID | ||
| F3 | 0,571 | 0,678 | 0,701 | 0,699 | 0,691 | ID | |
| F8 | 0,573 | 0,676 | 0,697 | 0,696 | 0,688 | 0,996 | ID |
Ghi chú: hoàn toàn giống nhau (ID)
Mức tương đồng về trình tự nucleotide của 7 chủng FAdV biến thiên rất rộng từ thấp nhất là 57,1% (chủng F3 so với chủng F5) cho đến cao nhất là 99,6% (chủng F3 so với chủng F8). Kết quả này dự đoán rằng 7 chủng virus được giải trình tự có thể thuộc về một số genotype FAdV khác nhau.
2.2. Kết quả phân tích trình tự amino acid của protein hexon
Trình tự aminio acid suy diễn của phân đoạn protein hexon (aa 121 đến aa 324) được trình bày ở hình 2.22.

Hình 2.22. Trình tự amino acid proteinhexon của 07 chủng FadV
Các vùng được đánh dấu A, B, C là những vùng ổn định, ít có sự thay đổi amino acid.
Đoạn gen hexon được giải trình tự ở nghiên cứu này (từ nucleotide 361 đến 972) mã hóa cho 204 codon (từ amino acid 121 đến 324). Trong 204 codon này (trừ 7 vị trí codon có đột biến mất nucleotide), có 52 codon trong đó sự thay đổi nucleotide ở bộ 3 mã hóa dẫn tới thay đổi amino acid được mã hóa (non-synonymous substitution), 145 codon còn lại có sự thay đổi nucleotide nhưng không dẫn tới thay đổi amino acid được mã hóa (synonymous substitution).
Có 3 vùng (A, B, C, hình 2.22) được xác định là những vùng ổn định về trình tự amino acid của protein hexon. Các phân tích sâu về tương quan giữa thay đổi nucleotide và amino acid tương ứng ở 3 vùng kể trên được tóm tắt ở hình 2.23.
Nhận thấy rằng, phần lớn các thay đổi nucleotide ở 3 vùng (A, B, C, hình 2.23) đều nằm ở vị trí thứ 3 của mỗi codon. Dựa vào bảng mã di truyền, người ta đã thấy rằng có khoảng 5%, 0% và 70% sự thay đổi nucleotide ở vị trí thứ 1, thứ 2 và thứ 3 của mỗi codon sẽ không dẫn tới sự thay đổi amino acid được mã hóa. Điều này lý giải vì sao trình tự nucleotide ở 3 vùng A, B, C kể trên của 7 chủng FAdV rất đa dạng, nhưng lại ít dẫn tới sự thay đổi amino acid được mã hóa.

Hình 2.23. Đặc điểm thay đổi codon ở một số vùng ổn định của gen hexon
Để dễ quan sát, mỗi nhóm gồm 3 nucleotide mã hóa cho amino acid tương ứng được nhóm và ngăn cách bởi vùng màu xám.
2.3. Kết quả phân tích đặc điểm dịch tễ học phân tử của FAdV
Trình tự gen mã hóa protein hexon từ amino acid 121 đến 324 đã được dùng để nghiên cứu đặc điểm dịch tễ học phân tử của các chủng FAdV phân lập tại Việt Nam và so sánh với các chủng virus lưu hành trên thế giới (hình 2.24).
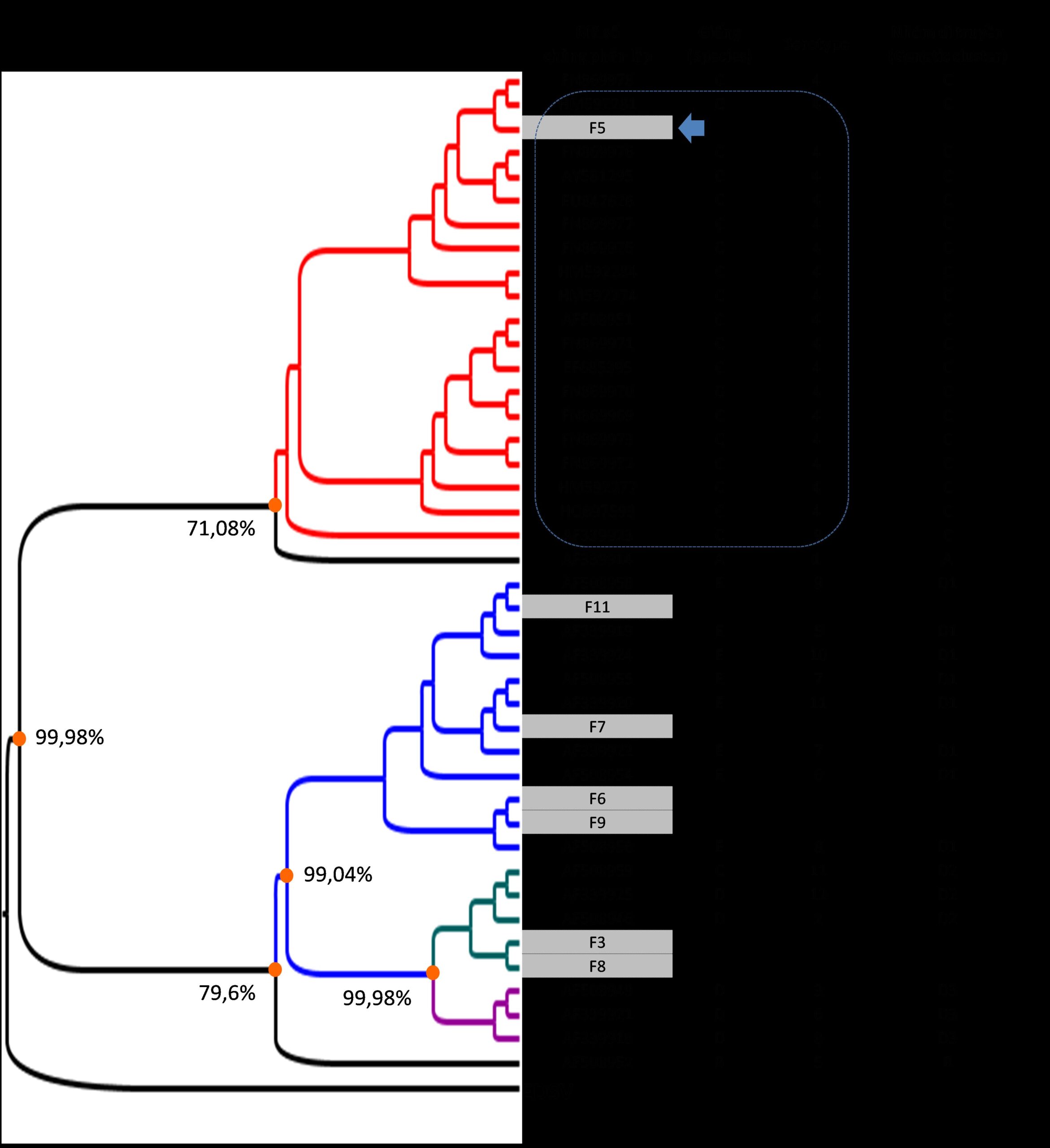
Hình 2.24. Cây phát sinh chủng loại của FadV
Ghi chú: các chủng FAdV với nhóm di truyền đã biết được dùng làm tham chiếu. 7 chủng phân lập trong nghiên cứu này được đánh dấu màu xám. Virus gây bệnh giảm đẻ ở gà (EDSV) được dùng làm outgroup.
Kết quả phân tích cây phát sinh chủng loại cho thấy 7 chủng FAdV phân lập đều thuộc về 3 giống là: FAdV-C (chủng phân lập F5), FAdV-D (F3, F8) và FAdV-E (chủng phân lập F6, F7, F9, F11). Không có chủng phân lập nào thuộc FAdV-A hoặc FAdV-B. Theo cách phân loại dựa vào trình tự gen (Meulemans & cs 2004), giống FAdV-D, FAdV-E được chia thành 3 nhóm di truyền là D1, D2 và D3. Với cách phân loại này, có 4 chủng (F6, F7, F9 và F11) thuộc về nhóm di truyền D1 và có 2 chủng (F3, F8) thuộc về nhóm di truyền D2. Với kết quả phân loại kể trên, có thể nhận thấy các chủng FAdV lưu hành ở khu vực lấy mẫu đa dạng về mặt di truyền.
Đối với FAdV, do sự phân loại virus dựa vào phản ứng huyết thanh học (serotype) và dựa vào đặc điểm gen (genotype) là khác nhau (Hess, 2000), vì vậy có 6/7 chủng phân lập được nghiên cứu chưa xác định được serotype. Đáng chú ý, có 1 chủng phân lập F5 được xếp vào nhóm di truyền FAdV-C và có quan hệ di truyền gần với các chủng FAdV thuộc serotype 4. Kết quả này cho phép suy diễn rằng chủng phân lập FAdV F5 thuộc serotype 4, serotype này được biết đến gây bệnh tích nước bao tim và viêm gan ở gà (Zhao & cs, 2015; Ye & cs, 2016).
Kết quả nghiên cứu đặc điểm dịch tễ học phân tử của FAdV serotype 4
Trong phần này, chúng tôi đi sâu tìm hiểu đặc điểm dịch tễ học phân tử của chủng FAdV4 thu được. Kết quả sàng lọc các trình tự gen tương đồng 99% với chủng F5 FAdV4 được trình bày ở hình 2.25.
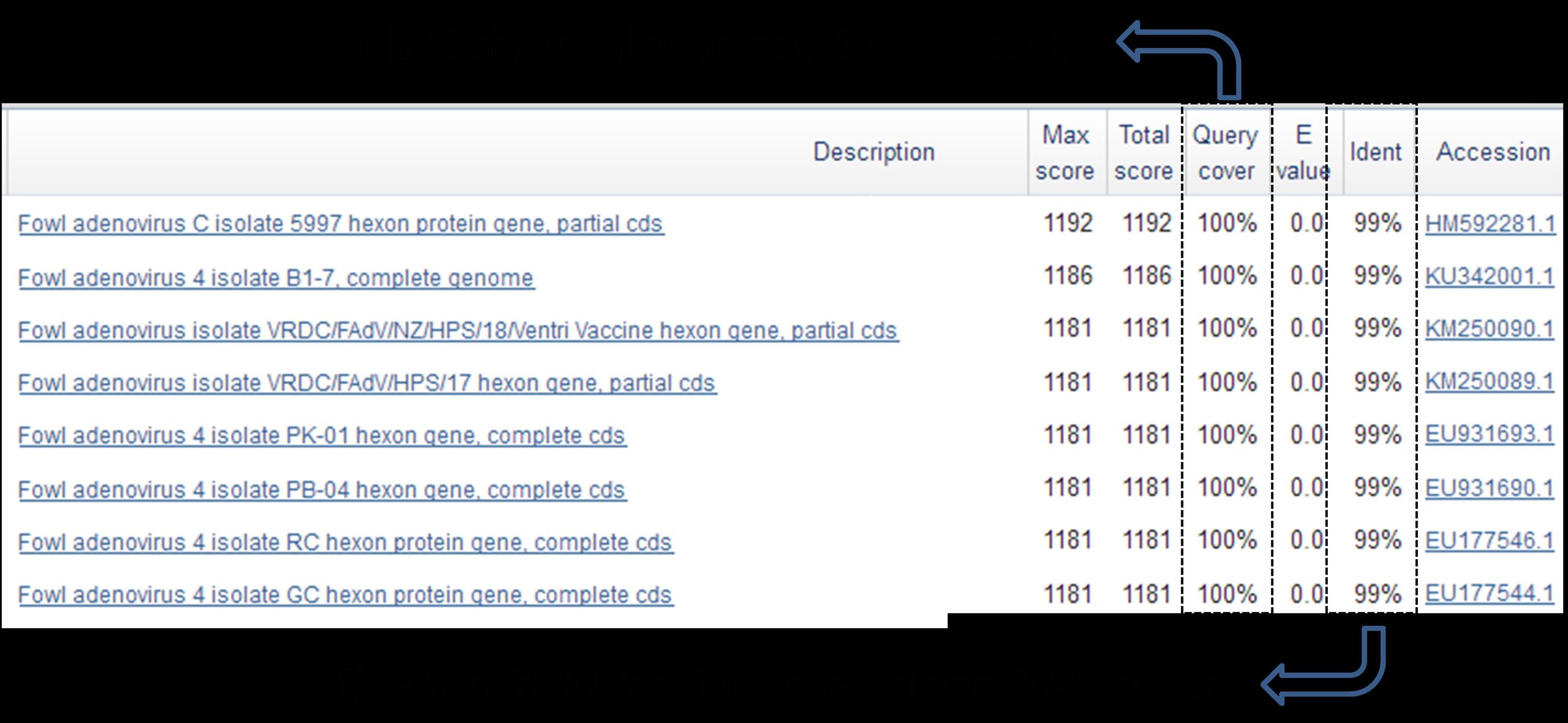
Hình 2.25. Kết quả BLAST tìm kiếm trình tự gen tương đồng 99% với chủng F5
Kết quả BLAST (minh họa một phần ở hình 2.25) cho 100 trình tự có mức tương đồng cao nhất (98%- 99%). Trong đó, chỉ có 71 chủng thoản mãn đầy đủ các điều kiện: (i) tương đồng ít nhất 99% với chủng F5 khi toàn bộ chiều dài của đoạn gen được so sánh, (ii) có đầy đủ thông tin về năm và địa điểm phân lập. Kết quả phân tích đặc điểm dịch tễ học phân tử theo không gian của serotype FAdV4 được trình bày ở hình 2.26.
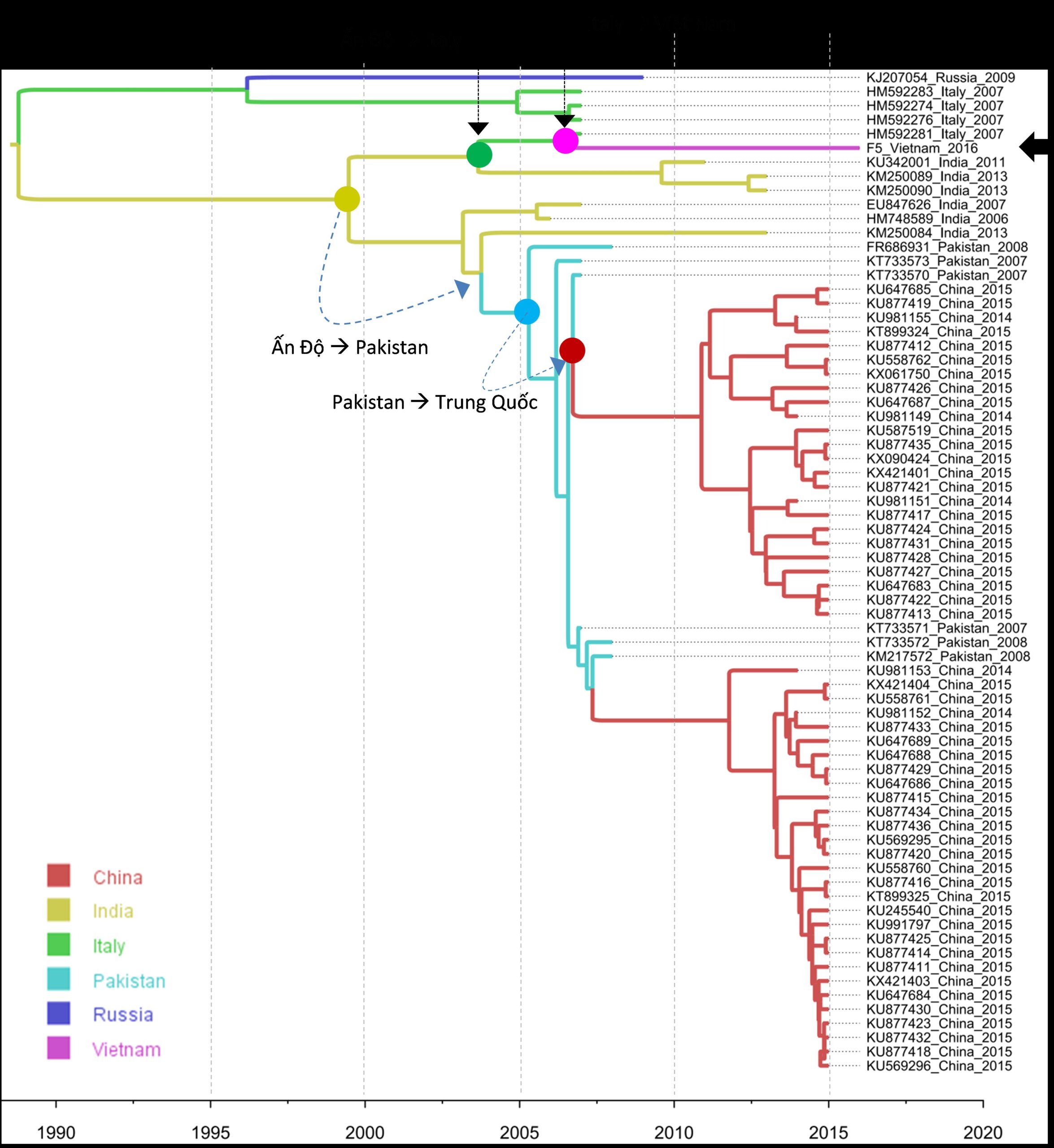
Hình 2.26. Đặc điểm dịch tễ học phân tử theo không gian của các chủng FAdV4
Ghi chú: phân tích này chỉ được thực hiện với các chủng FAdV4 có đặc điểm giống ít nhất 99% với chủng F5 lưu hành ở Việt Nam.
Các chủng FAdV4 có trình tự gen giống 99% với chủng F5 của Việt Nam được thấy ở một số nước thuộc châu Âu (Nga, Italy) và châu Á (Trung Quốc, Ấn Độ, Pakistan). Kết quả cho biết FAdV4 lưu hành ở các nước châu Á có thể có nguồn gốc từ Ấn Độ, với con đường truyền lây được dự đoán như sau: (i) với các nhánh virus lưu hành ở Trung Quốc có nguồn gốc từ Ấn Độ -> Pakistan -> Trung Quốc; (ii) với nhánh virus lưu hành ở Việt Nam có nguồn gốc từ Ấn Độ -> Italy -> Việt Nam. Với kết quả trên có thể thấy rõ rằng chủng F5 FAdV4 lưu hành ở Việt Nam nằm ở nhánh di truyền khác với các chủng cùng loại lưu hành ở Trung Quốc. Phân tích này cũng dự đoán thời điểm FAdV4 xuất hiện ở nước ta sớm nhất vào khoảng năm 2005 (hình 2.26).
Đã có một số nghiên cứu trên thế giới đã phân tích đặc điểm dịch tễ học phân tử của FAdV lưu hành ở các nước khu vực châu Âu (Niczyporuk, 2016), châu Á (Mase & Nakamura, 2014). Nhưng theo hiểu biết của chúng tôi, chưa có nghiên cứu nào đề cập đến đặc điểm dịch tễ học phân tử của virus theo không gian và thời gian để chỉ rõ nguồn gốc của các chủng virus lưu hành ở một quốc gia nhất định. Với kết quả thu được trong nghiên cứu này gợi mở hướng nghiên cứu sâu hơn về đặc điểm dịch tễ học phân tử của FAdV nói chung và của các serotype FAdV gây bệnh nói riêng.
Điều cần lưu ý là mặc dù đã phát hiện nhiều genotype virus, nhưng số chủng thuộc genotype gây bệnh (FAdV4) là ít so với các chủng phát hiện được; do vậy cần lưu ý là phát hiện adenovirus nói chung không đồng nghĩa với bệnh (vì chỉ có 1 số serotype/genotype được chứng minh là gây bệnh).
Tài liệu tham khảo
Ganesh, K., Suryanarayana, V. V. S. & Raghavan, R. (2002). “Detection of fowl adenovirus associated with hydropericardium hepatitis syndrome by a polymerase chain reaction.” Veterinary Research Communications 26(1): 73-80.
Lê Văn Trường Huỳnh, Thị Mỹ Lệ, Nguyễn Văn Giáp (2020). Nghiên cứu sự lưu hành của Fowl Adenovirus ở gà nuôi tại Hà Nội và vùng phụ cận. Tạp chí khoa học nông nghiệp Việt Nam, 18 (8): 588-598.

 English
English