Đỗ Thơm1,2, Bùi Khánh Linh2, Rika Umemiya-Shirafuji1, Taiwan Inpankaew3, Tanjila Hasan1, Iqra Zafar1, Zhuongwei Ma1, Li Hang1, Uday Kumar Mohanta1, Moaz Amer1, Shimaa Abd El-Salam El-Sayes1,4, Xuenan Xuan1*,
Ketsarin Kamyingkird3*.
1National Research Center for Protozoan Diseases, Obihiro University of Agriculture and Veterinary Medicine, Obihiro, Hokkaido, Japan
2Department of Parasitology, Faculty of Veterinary Medicine, Vietnam National University of Agriculture, Hanoi, Vietnam
3Department of Parasitology, Faculty of Veterinary Medicine, Kasetsart University, Bangkok, Thailand
4Department of Biochemistry and Molecular Biology, Faculty of Veterinary Medicine, Mansoura University, Mansoura, Egypt
Tính cấp thiết
Những người nuôi chó rất lo lắng về sự lây nhiễm của bọ ve trong vật nuôi của họ. Các tỷ lệ phổ biến và sự lây nhiễm của bọ ve và vi sinh vật gây bệnh của chúng đã bị hạn chế theo quan điểm của những người nuôi chó ở Việt Nam. Nghiên cứu này điều tra sự hiện diện của sự lây nhiễm của bọ ve và các mầm bệnh liên quan với nó ở trên chó trong những được đưa đến các bệnh viện thú y ở Việt Nam. Trong khảo sát, 1.423 đã tham gia từ tháng 2 đến tháng 10 năm 2022. Các phương pháp phân tử và hình thái được sử dụng để xác định bọ ve và các mầm bệnh. Ngoài ra, các biến rủi ro liên quan đến sự lây nhiễm của bọ ve đã được ghi nhận và phân tích bằng các phương pháp thống kê. Tổng mức độ tiếp xúc với ve chó nâu (Rhipicephalus sanguineus sensu lato) là 29,01%. Nam Định tỷ lệ mắc bọ ve cao nhất trong các khu vực nghiên cứu. Sự lây nhiễm của bọ ve đạt mức cao đỉnh điểm từ tháng 6 đến tháng 9 ở khu vực phía bắc của đất nước, với các mùa riêng biệt cho thấy mối tương quan chặt chẽ với sự lây nhiễm của bọ ve ở chó. Trong số 177 mẫu bọ ve được kiểm tra, 146 (82,49%) có kết quả xét nghiệm dương tính với ít nhất một loại nhiễm trùng. Mycoplasma spp. (78,53%) là phổ biến nhất, tiếp theo là Anaplasma spp. (37,29%), Rickettsia felis (5,08%), Babesia vogeli và Hepatozoon canis (2.82%). Trong nghiên cứu hiện tại, có một mối liên hệ thống kê có ý nghĩa giữa sự lây nhiễm của bọ ve và các đặc điểm như tuổi, giống, kích thước cơ thể, lối sống và tần suất tắm. Hiểu vè vec tơ truyền bệnh theo mùa của bọ ve là rất quan trọng để xác định các cá thể hoặc động vật dễ mắc các bệnh do ve truyền. Nghiên cứu sự phân bố của bọ ve và khả năng mang và lan truyền các ví sinh vật zoonotic từ động vật sang người ở những nơi cụ thể có thể hỗ trợ bác sĩ thú y và các nhà hoạch định chính sách trong việc thực hiện các chiến lược hiệu quả để kiểm soát nhiễm vi sinh vật zoonotic từ động vật sang người.
Mục đích nghiên cứu
Nghiên cứu này điều tra sự hiện diện của nhiễm ve và các tác nhân gây bệnh liên quan ở chó nuôi tại các bệnh viện thú y ở Việt Nam.
Phương pháp nghiên cứu:
2.1 Sự đồng ý về mặt đạo đức:
Ủy ban Đạo đức của Đại học Nông nghiệp và Thú y Obihiro đã chấp nhận quy trình sử dụng mẫu động vật trong nghiên cứu này (Giấy phép thí nghiệm trên động vật: 21-25; Thí nghiệm DNA: 1725-5).
2.2 Khu vực nghiên cứu, thu thập mẫu và xác định ve
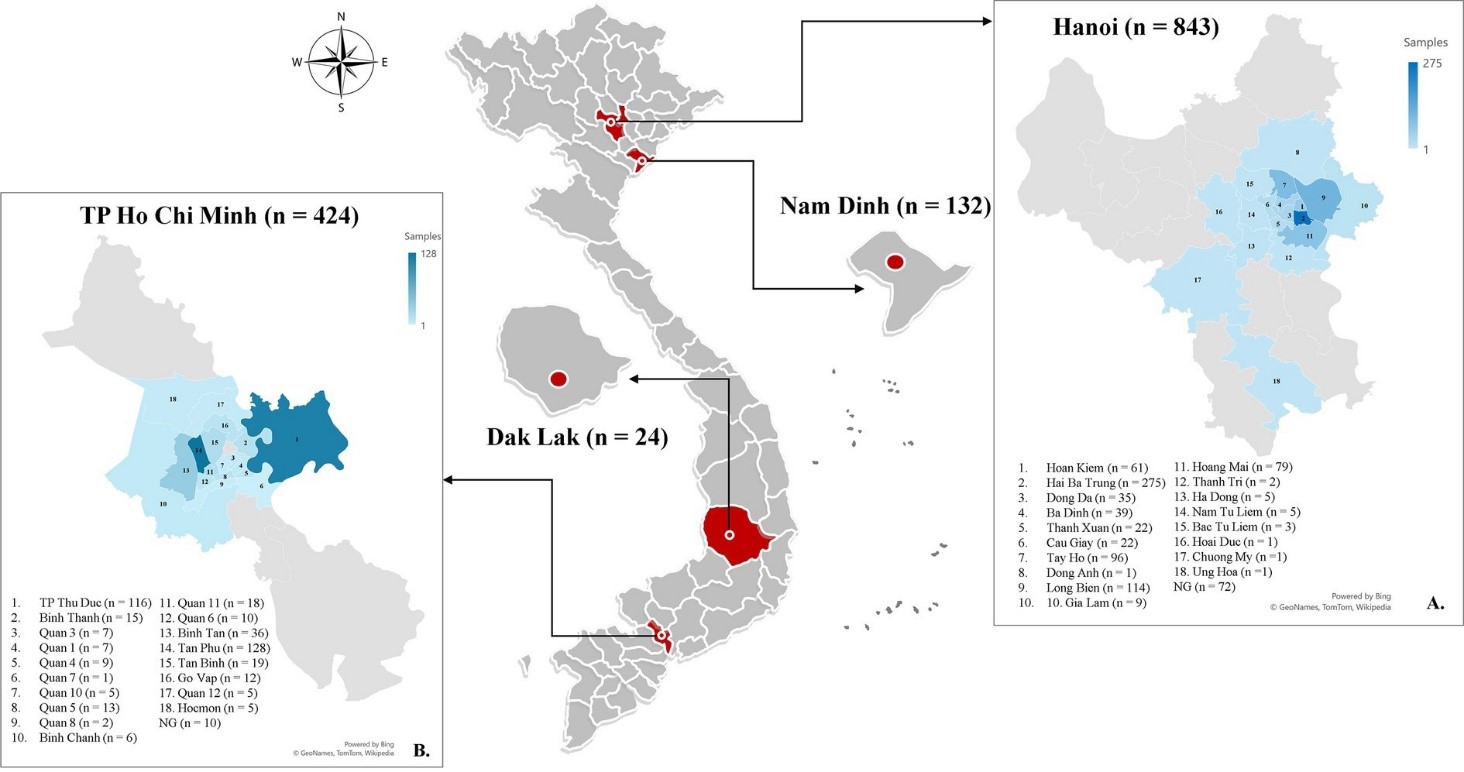
Một cuộc khảo sát về nhiễm ve ở chó đã được thực hiện từ tháng 2 đến tháng 10 năm 2022 với sự tham gia tự nguyện từ các phòng khám thú y ở Hà Nội (21°01′N, 105°51′E), Thành phố Hồ Chí Minh (10°46′N; 106°42′E), Nam Định (20°25′N; 106°10′E) và Đắk Lắk (12°4′N, 108°3′E) tại Việt Nam (Hình 1). Các bác sĩ thú y đã tiến hành kiểm tra toàn diện các động vật để phát hiện ve bằng cách kiểm tra toàn bộ bề mặt cơ thể trong hơn 5 phút. Các cuộc kiểm tra bao gồm đánh giá tình trạng thể chất của động vật và ghi chú bất kỳ bất thường nào trên da/lông và màng nhầy như vàng da, nhợt nhạt hoặc chảy máu. Một bảng câu hỏi đã được tạo ra để thu thập dữ liệu về vị trí, giống, giới tính, tuổi và kích thước cơ thể của các chó được lấy mẫu nhằm phân tích một số danh mục liên quan đến nhiễm ve. Các câu hỏi bổ sung được đưa ra về môi trường sống (trong nhà, ngoài trời hoặc bán ngoài trời) và tần suất tắm (hàng tuần, hàng tháng, hai tháng một lần hoặc hàng năm) nếu có.
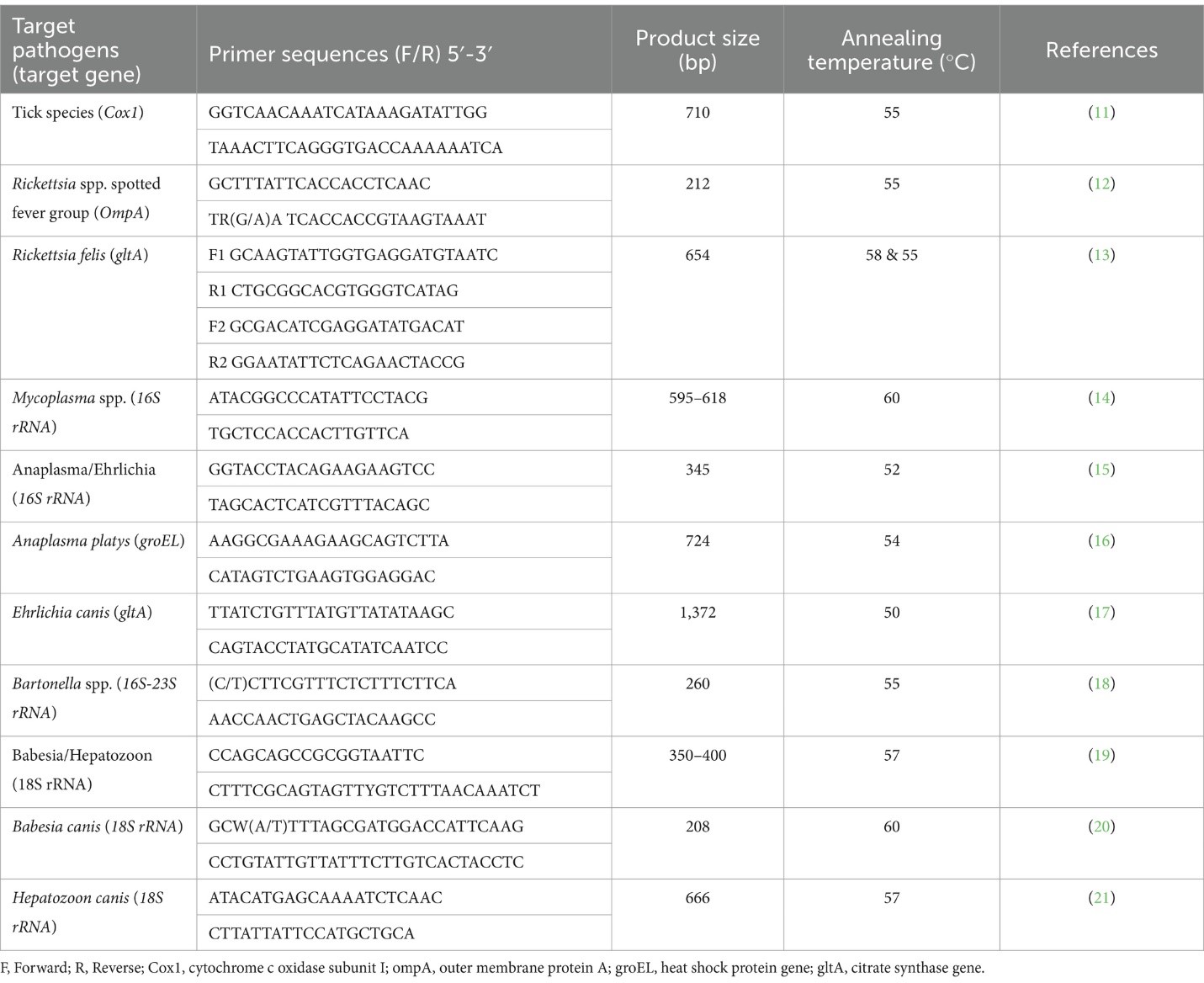
Từ một đến năm con ve gắn trên chó đã được thu thập và lưu trữ trong các ống 1.5 mL chứa 70% ethanol. Các mẫu vật sau đó được chuyển đến Trung tâm Nghiên cứu Các bệnh do Protozoa ở Obihiro, Hokkaido, Nhật Bản để xác định bằng phân tích hình thái và phân tử. Tổng cộng có 555 con ve đã được gửi đi để xác định loài và phát hiện vi sinh vật. Các con ve được kiểm tra bằng kính hiển vi soi nổi (Olympus SZX16) để xác định loài và giai đoạn sống (ấu trùng, nhộng, hoặc trưởng thành) bằng cách sử dụng các đặc điểm hình thái cụ thể. Sau khi xác định hình thái ban đầu, các con ve được rửa sạch bằng dung dịch đệm phosphate. Sau đó, các con ve cùng giai đoạn sống và thu thập từ cùng một con chó được gộp lại để chiết xuất DNA bằng bộ DNeasy Blood and Tissue Kits (Quiagen, Đức) theo hướng dẫn của nhà sản xuất. Sau quá trình gộp mẫu, tổng cộng có 177 mẫu gộp được thu được, với mỗi mẫu gộp chứa từ một đến năm con ve riêng lẻ để chiết xuất DNA. Sau khi chiết xuất DNA, các trình tự nhắm vào gen đặc hiệu cho loài đã được khuếch đại để xác nhận phân tử loài ve và các vi sinh vật liên quan.(Bảng 1)
2.3 Phát hiện phân tử và phân tích hệ gen của vi sinh vật
Các mẫu DNA đã được sàng lọc để phát hiện các protozoa apicomplexan (như Babesia spp., Hepatozoon spp.), vi khuẩn thuộc họ Anaplasmataceae (như A. platys, E. canis), Mycoplasma spp., Rickettsia spp., và Bartonella spp. bằng cách sử dụng các bộ mồi thông qua phản ứng PCR thông thường (cPCR). Phản ứng cPCR đã khuếch đại các trình tự mục tiêu của các loài Ehrlichia/Anaplasma spp. (gen 16S rRNA), Babesia/Hepatozoon spp. (gen 18S rRNA), và Rickettsia spp. từ nhóm sốt phát ban (OmpA). Các mẫu dương tính cho chi của các loài đã nêu sau đó được phân tích bằng cách sử dụng các mồi nhắm vào các trình tự đặc hiệu cho loài thông qua cPCR hoặc nested PCR. Tất cả các mồi và các gen mục tiêu được sử dụng để phát hiện tác nhân gây bệnh đã được trình bày trong Bảng 1. Các mẫu dương tính với tác nhân gây bệnh và nước cất đã được sử dụng làm đối chứng dương tính và âm tính, tương ứng, cho tất cả các thí nghiệm. Các sản phẩm PCR được kiểm tra bằng cách điện di trên gel agarose 1.5% (LE agarose, Thermo Fisher Scientific, Waltham, MA, Hoa Kỳ) và được quan sát dưới đèn huỳnh quang UV (ATTO, Tokyo, Nhật Bản). DNA đã được chiết xuất và tinh chế từ sản phẩm dương tính bằng cách cắt nó ra khỏi gel sử dụng bộ chiết xuất gel từ QIAGEN, Đức. Kết quả, đã được tinh chế và có các trình tự mong muốn, đã trải qua quá trình giải trình tự bằng bộ BigDye v3.1 Terminator Cycle Sequencing Kit trên máy phân tích gen ABI PRISM 3100 của Applied Biosystem, Hoa Kỳ.
Các biểu đồ trình tự đã được đánh giá bằng phần mềm BioEdit (phiên bản 7.5.2) và so sánh với các trình tự trong cơ sở dữ liệu Genbank bằng cách sử dụng Công cụ Tìm kiếm Căn chỉnh Địa phương Cơ bản (BLAST) trên trang web của Trung tâm Thông tin Công nghệ Sinh học Quốc gia Hoa Kỳ (NCBI). Công việc này đã đánh giá sự biến đổi di truyền của Mycoplasma spp. và Rickettsia felis bằng cách kiểm tra các trình tự dựa trên gen 16S rRNA của Mycoplasma spp. (618 bp) và các trình tự dựa trên gen gltA của R. felis (654 bp) bằng cách sử dụng phân tích hệ gen với công cụ MEGA X. Các cây hệ gen đã được tạo ra bằng phương pháp tối đa hóa khả năng và mô hình thay thế phù hợp nhất. Phân tích bootstrap với 1,000 lần lặp lại đã được thực hiện để đánh giá độ tin cậy của các mẫu nhánh trong các cây. Mỗi trình tự đều được cung cấp số truy cập Genbank, nguồn gốc cách ly và quốc gia xuất xứ.
2.4 Phân tích thống kê
Ứng dụng OpenEpi đã được sử dụng để tính toán tỷ lệ phát hiện nhiễm ve, xác định tỷ lệ phần trăm các bệnh được phát hiện và ước lượng khoảng tin cậy 95% (95% CI). Một bài kiểm tra chi-squared đã được sử dụng để điều tra mối quan hệ thống kê giữa tỷ lệ nhiễm ve và các yếu tố độc lập của vật chủ. Tỷ lệ odds (OR) đã được tính toán để đánh giá mức độ mạnh mẽ của mối quan hệ giữa từng danh mục và nhiễm ve, xác định xem có sự liên kết có ý nghĩa nào giữa chúng hay không. Mức độ ý nghĩa thống kê được thiết lập ở mức p ≤ 0.05. Nghiên cứu đã sử dụng mức độ tin cậy 95%. Công cụ Statulator đã được sử dụng để phân tích dữ liệu.
Kết quả chính:
3.1 Dữ liệu mẫu và xác định hình thái ve
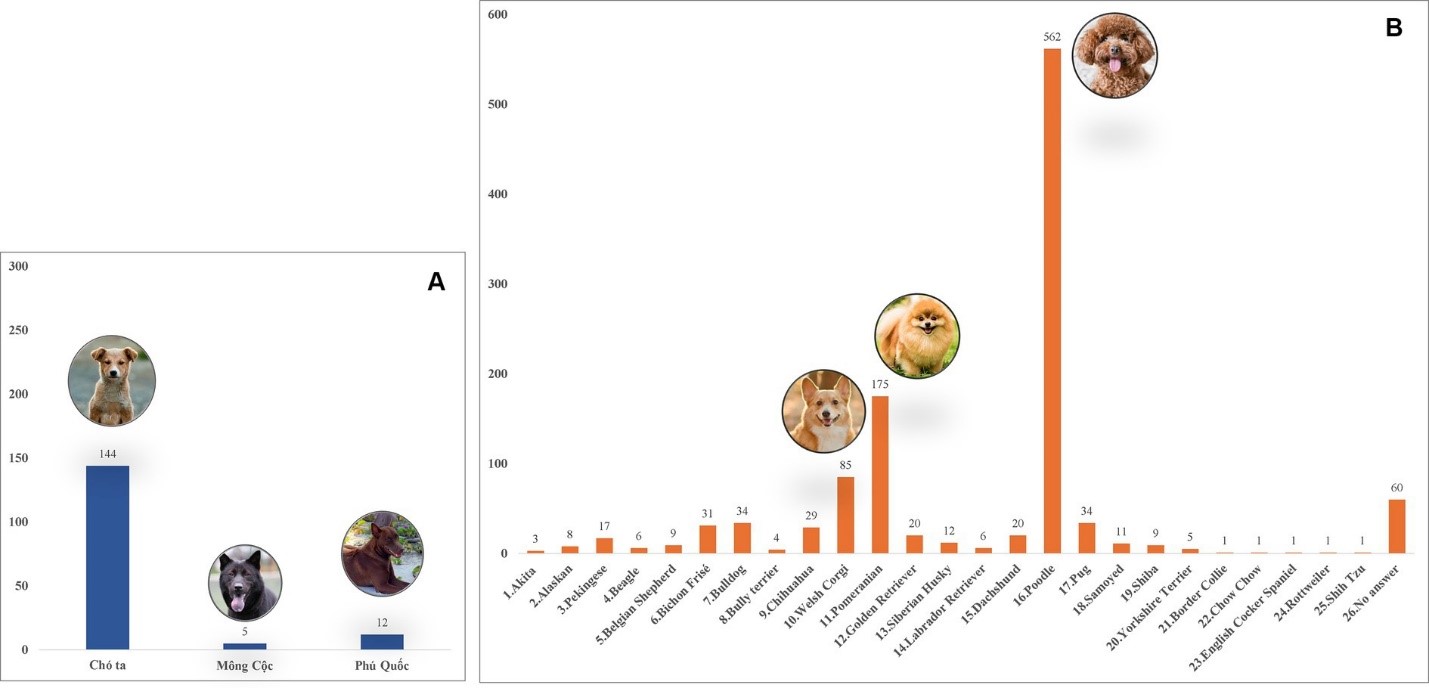
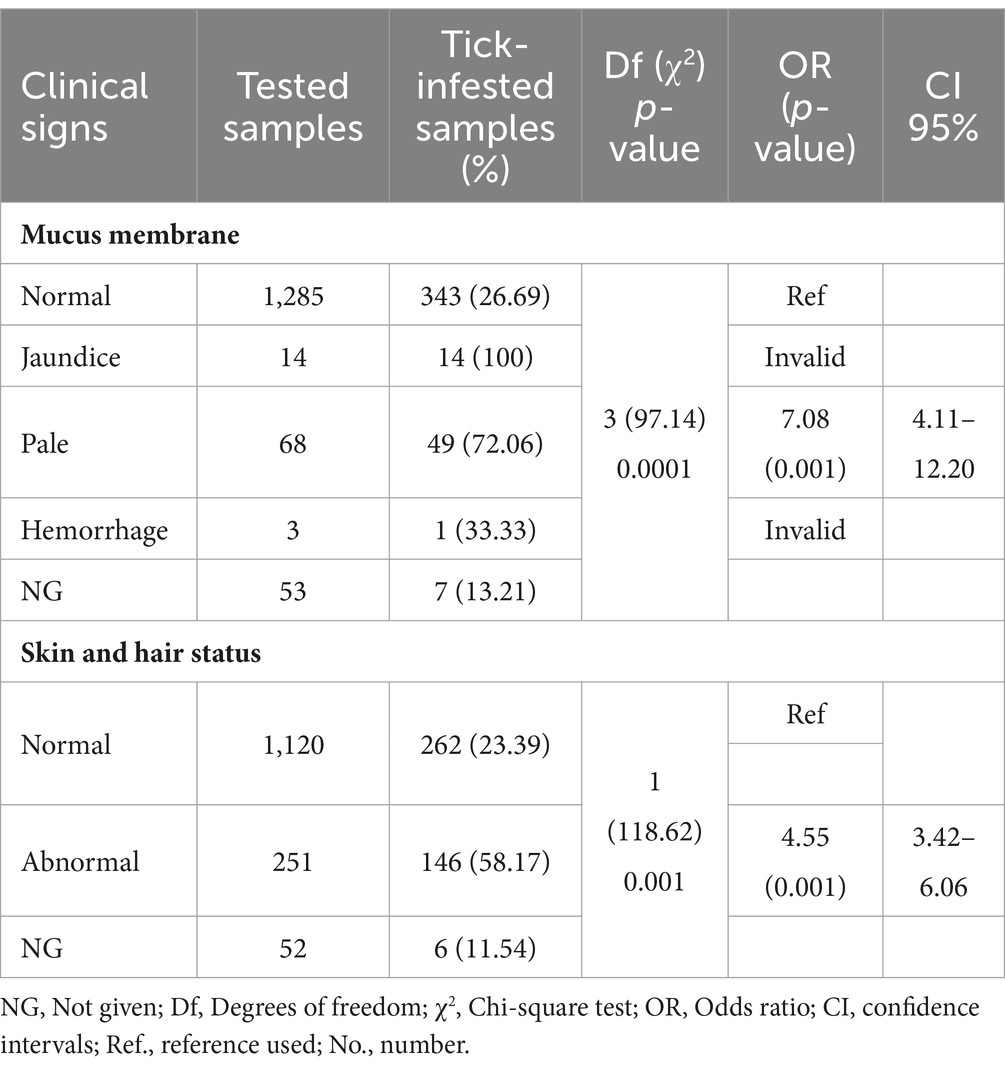
Tổng cộng có 1,423 con chó tham gia khảo sát, trong đó 843 con từ Hà Nội (59.24%), 424 con từ Thành phố Hồ Chí Minh (29.8%), 132 con từ Nam Định (9.28%), và 24 con từ Đắk Lắk (1.68%; Hình 1). Các con chó được ghi danh bao gồm: 630 con cái (44.27%), 729 con đực (51.23%), và 64 con có dữ liệu không được báo cáo (4.5%). Độ tuổi của các con vật dao động từ 2 tháng đến 20 năm, với khoảng 43.5% (619) trong tổng số chó nằm trong độ tuổi từ 1 đến 5 năm. Các con chó thuộc hơn 20 giống, được chia thành ba nhóm. Chi tiết về các giống chó khác nhau được quan sát trong nghiên cứu hiện tại được trình bày trong (Hình 2); trong đó, 1,145 con (80.46%) là giống ngoại, 161 con (11.31%) là giống nội. Phần lớn sống trong nhà (662, 46.52%), kích thước nhỏ (≤3.5 kg; 462, 32.46%), và được tắm ít nhất một lần mỗi tuần (707, 49.68%). Các triệu chứng lâm sàng như bất thường ở da và màng nhầy được phát hiện có liên quan thống kê đến sự hiện diện của nhiễm ve ở chó (Bảng A1). Tổng cộng có 555 con ve đã được gửi đi để xác định loài và phát hiện vi sinh vật. Cụ thể, dựa trên các đặc điểm hình thái điển hình, 33 ấu trùng, 75 con nhộng, và 447 con trưởng thành (195 con đực và 252 con cái) đã được xác định thuộc về phức hợp loài R. sanguineus s.l.(Hình 3)

3.2 Phân bố địa lý và phân bố theo mùa của ve
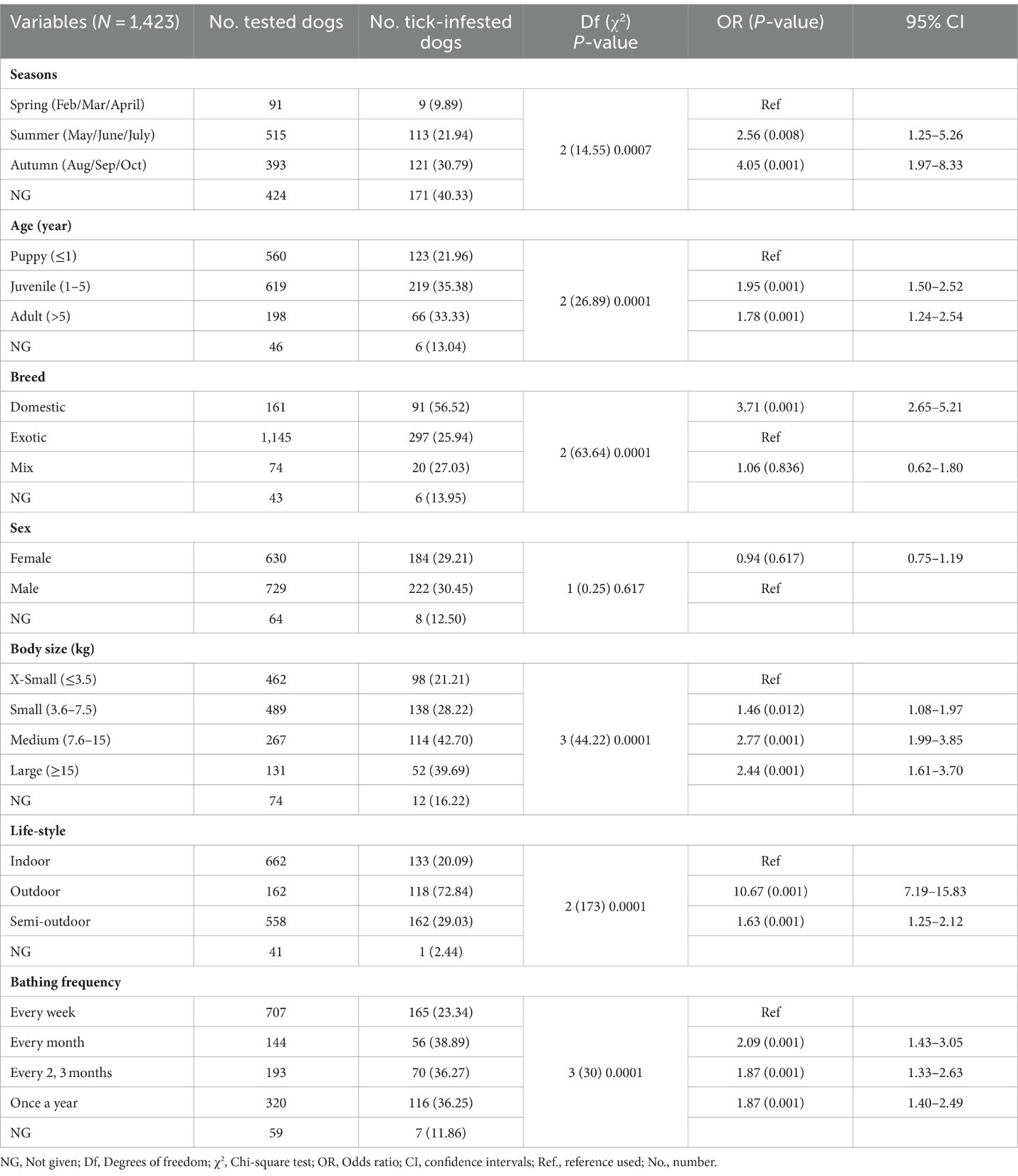
Trong số 1,423 con chó được kiểm tra, có 414 con (29.01%, 95% CI: 26.79–31.51%) mang ít nhất một con ve tại thời điểm lấy mẫu. Theo phân tích thống kê, có sự khác biệt có ý nghĩa giữa các khu vực địa lý và tỷ lệ nhiễm ve ở chó. Tỷ lệ nhiễm ve cao nhất ở chó được lấy mẫu tại Nam Định với 55.3% (73/132, 95% CI: 46.79–63.52), tiếp theo là Thành phố Hồ Chí Minh với 39.86% (169/424, 95% CI: 35.31–44.59) và Hà Nội với 20.7% (170/843, 95% CI: 17.6–23.01). Tại Đắk Lắk, sự hiện diện của ve được phát hiện ở hai con chó (8.33%, 95% CI: 2.3–25.85). Ngoài ra, chó sống ở Nam Định và Thành phố Hồ Chí Minh có khả năng nhiễm ve cao gấp 4.9 lần (p < 0.001, 95% CI: 3.34–7.18%) và 2.62 lần (p < 0.001, 95% CI: 2.03–3.39%) so với những con ở Hà Nội. Các mẫu thu thập từ miền Bắc Việt Nam đã được phân tích phân bố theo mùa của ve. Trong các mùa khác nhau, nhiều chó bị nhiễm ve từ tháng 5 đến tháng 9 và số lượng chó bị nhiễm ve giảm xuống từ tháng 2 đến tháng 4 (Hình 4). Thêm vào đó, tỷ lệ phát hiện nhiễm ve ở chó có liên quan thống kê với các mùa khác nhau ở miền Bắc. Cụ thể, phần lớn chó bị nhiễm ve trong nghiên cứu được phát hiện trong các tháng ấm áp của mùa hè (21.94%, 113/515) và mùa thu (30.79%, 121/393) so với mùa xuân (9.89%, 9/91). Trong mùa hè và mùa thu, chó có khả năng nhiễm ve cao gấp 2.56 (p = 0.008) và 4.05 lần (p < 0.001) so với những con trong mùa xuân, tương ứng (Bảng 2).
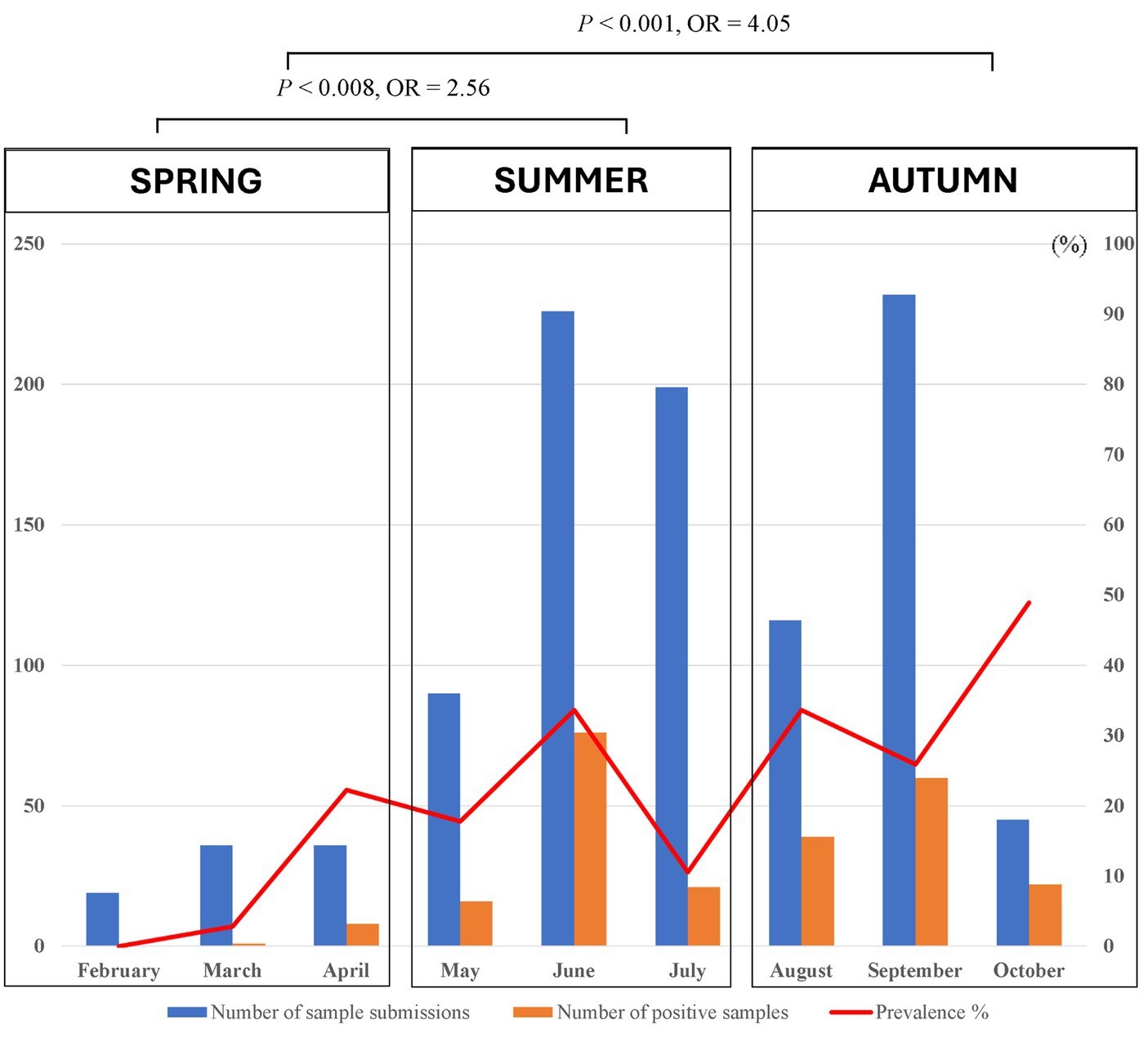
3.3 Các yếu tố rủi ro liên quan đến nhiễm ve ở chó tại Việt Nam
Mối quan hệ giữa nhiễm ve và các đặc điểm khác nhau của vật chủ đã được đánh giá thống kê trong nghiên cứu hiện tại và được báo cáo trong Bảng 2 cùng với số lượng và tỷ lệ phần trăm nhiễm ve. Các mẫu có thông tin thiếu đã bị loại khỏi phân tích. Phần lớn các con chó bị nhiễm ve được phát hiện là chó con (35.38%, 219/619), giống nội (56.52%, 91/161), kích thước trung bình (42.7%, 114/267), lối sống ngoài trời (72.84%, 118/162) và được tắm ít hơn một lần mỗi tháng (36.83%, 242/657), so với các nhóm khác trong cùng danh mục. Không có mối liên hệ có ý nghĩa thống kê giữa sự dương tính với ve và giới tính. Các con chó con và chó trưởng thành có khả năng nhiễm ve cao gấp 1.95 lần (p < 0.001) và 1.78 lần (p < 0.001) so với chó con, tương ứng. Chó nội có khả năng nhiễm ve cao gấp 3.71 lần so với chó ngoại (p = 0.001, 95% CI: 2.65–5.21). Chó có kích thước nhỏ có khả năng nhiễm ve thấp hơn gấp 2 lần so với chó kích thước trung bình (p = 0.001, 95% CI: 1.99–3.85) hoặc lớn (p = 0.001, 95% CI: 1.61–3.7). Chó có lối sống ngoài trời có mối liên hệ đáng kể với việc phát hiện ve và có nguy cơ nhiễm ve cao gấp 10 lần so với những con sống trong nhà trong nghiên cứu này (p < 0.001, 95% CI: 7.19–15.83).
3.4 Phát hiện vi sinh vật trong các mẫu ve thu thập từ chó ở Việt Nam
Trong số 177 mẫu ve được kiểm tra, có 146 mẫu (82.49%) phát hiện ít nhất một tác nhân gây bệnh, với Mycoplasma spp. là phổ biến nhất (78.53%, CI: 71.91–83.94), tiếp theo là Anaplasma spp. (37.29%, CI: 30.51–44.61), R. felis (5.08%, CI: 2.69–9.38), B. vogeli và H. canis (2.82%, CI: 1.04–6.82). Trong 79 mẫu, một tác nhân duy nhất được phát hiện (44.63%, CI: 37.5–51.99). Nhiều tác nhân được phát hiện trong 67 mẫu (37.85%, CI: 31.04–45.18; Bảng 3). Về các giai đoạn của ve, trong số 151 mẫu ve trưởng thành, có 121 mẫu (80.13%) có nhiễm Mycoplasma spp. Các mẫu này tiếp theo có nhiễm Anaplasma spp. (39.07%, 59/151), R. felis (5.3%, 8/151), B. vogeli (2.65%, 4/151), và H. canis (0.66%, 1/151). Trong cuộc điều tra này, các mẫu nymph và trưởng thành được phát hiện có cùng các nhiễm trùng, trong khi ấu trùng chủ yếu có Mycoplasma spp. (87.5%; Bảng 4).
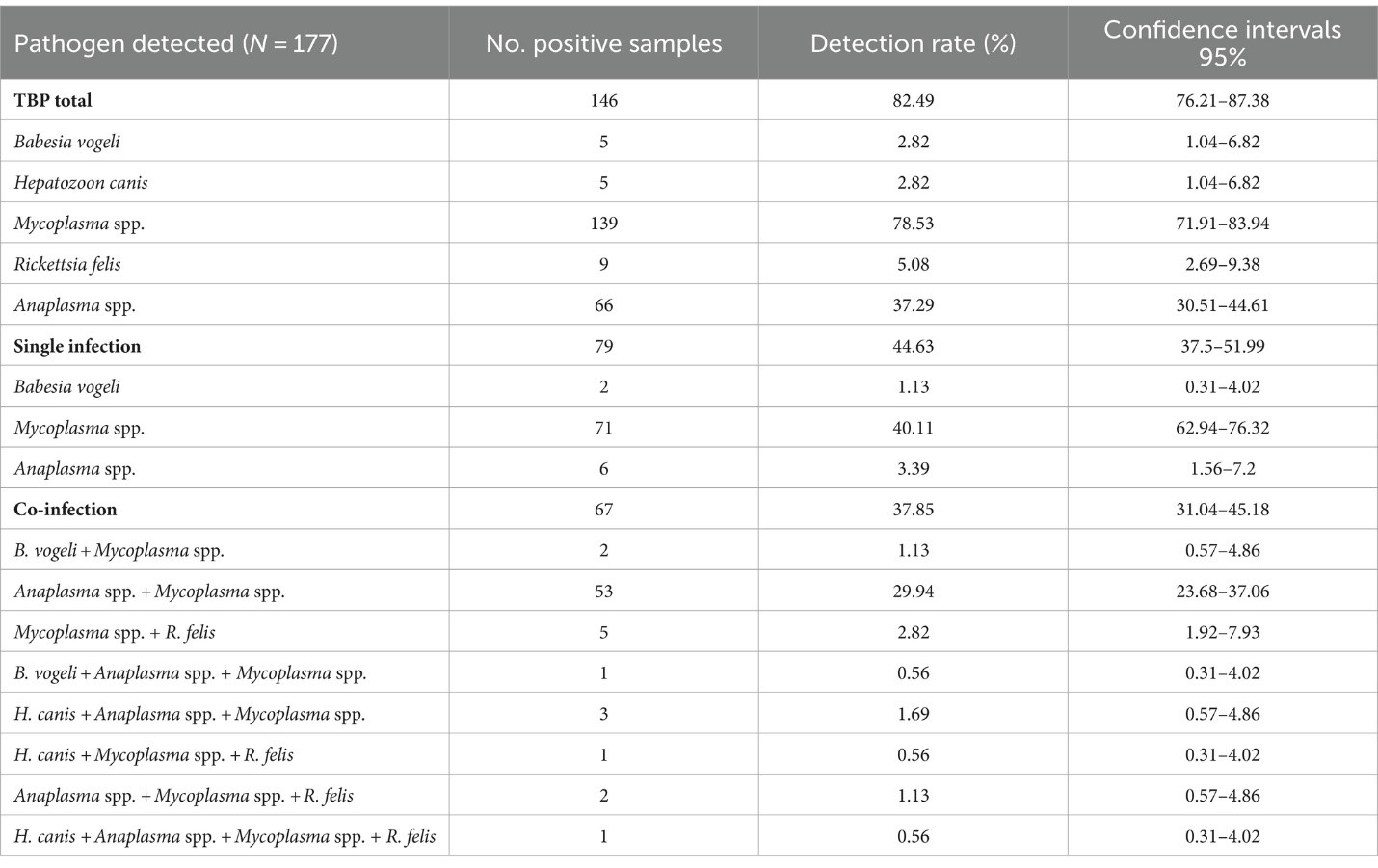
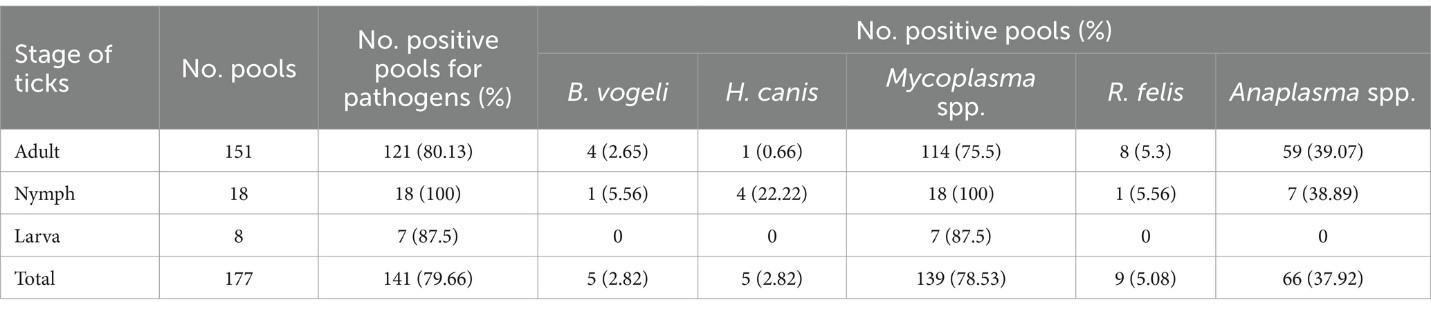
3.5 Giải trình tự danh tính và phân tích hệ gen
Các trình tự nucleotide của ve và mỗi tác nhân gây bệnh đã được xác định cho thấy độ tương đồng từ 99–100% với các trình tự trong cơ sở dữ liệu Genbank. Các mẫu của R. sanguineus s.l. đã được xác định gen là một phần của dòng nhiệt đới với độ tương đồng nucleotide 100% (số truy cập GenBank MF425992–MF425994). Các trình tự gen 18S rRNA của B. vogeli khớp với MT386936 với độ tương đồng 100%, trong khi H. canis cho thấy độ tương đồng 99.85% với MG050161. Các trình tự gen gltA của R. felis cho thấy mức độ tương đồng cao, dao động từ 99.84% đến 100%, với các trình tự OM936910–MT019627. Các trình tự gen 16S rRNA của Mycoplasma spp. cho thấy độ tương đồng nucleotide từ 99.51% đến 99.68% với các trình tự của Mycoplasma wenyonii (MF377464–MG948627). Trong khi đó, các trình tự gen 16S rRNA của Anaplasma spp. cho thấy mức độ tương đồng cao, dao động từ 99.03% đến 99.67%, với các trình tự của các loài khác nhau trong chi Anaplasma và Ehrlichia (EU090184–MN481611). Tuy nhiên, không có mẫu dương tính với Anaplasma/Ehrlichia nào cho kết quả dương tính với các xét nghiệm E. canis hoặc A. platys.
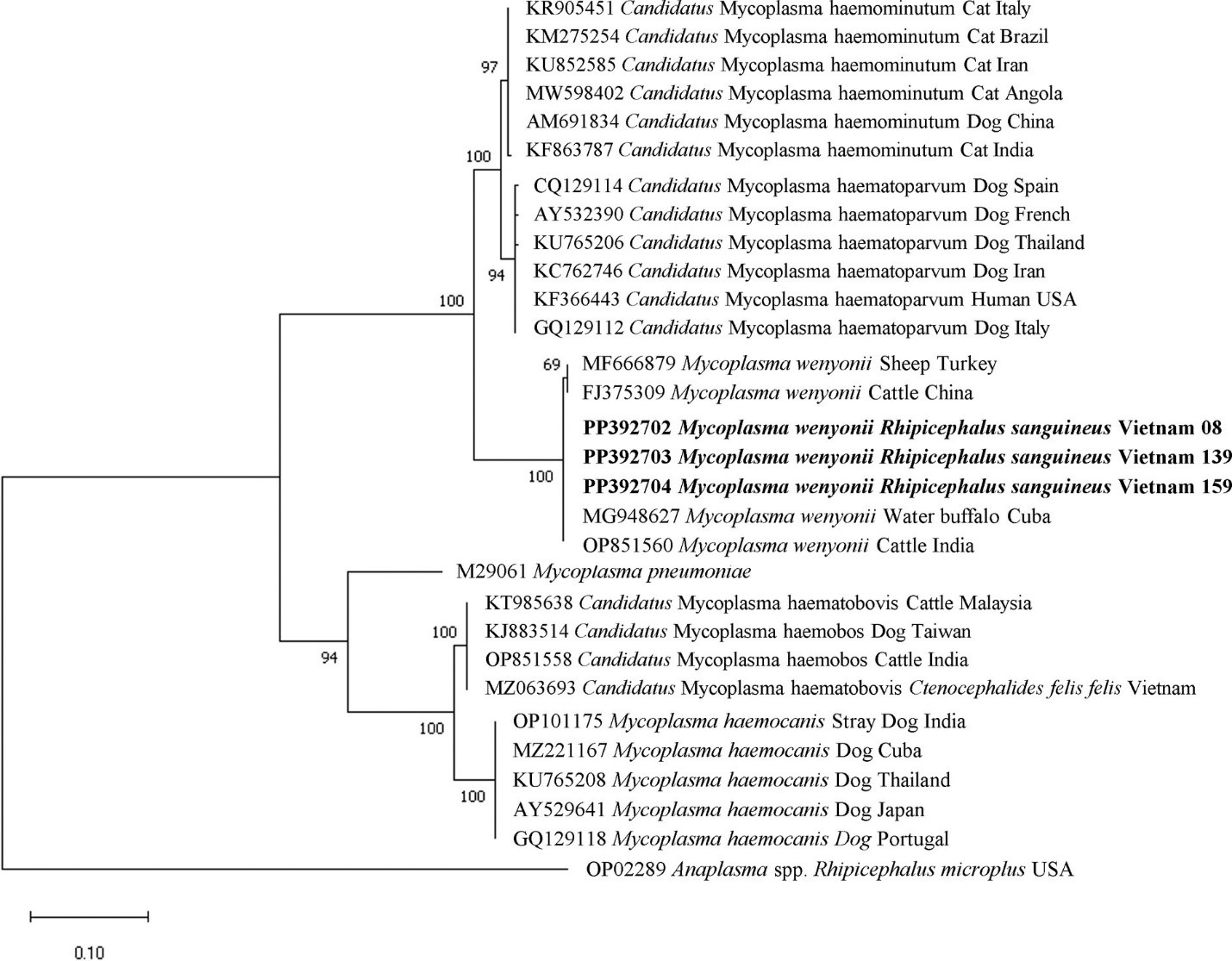
Việc xác định phân tử các trình tự cụ thể cho Mycoplasma spp. và R. felis đã được xác nhận bằng sự phân tách rõ ràng của các nhóm đặc hiệu cho loài thông qua phân tích hệ gen. Cây hệ gen dựa trên gen 16S rRNA của Mycoplasma spp. chỉ ra rằng tất cả các trình tự M. wenyonii (dài 618 bp) được nhóm lại trong một nhánh riêng biệt với sự hỗ trợ mạnh mẽ (giá trị bootstrap 99%). Nhánh này bao gồm các trình tự từ cùng một loài được tìm thấy ở nhiều vị trí địa lý khác nhau, trong khi loại trừ các loài Mycoplasma spp. khác (Hình 5). Phân tích hệ gen của các trình tự gen gltA không hoàn chỉnh (dài 654 bp) cho thấy tất cả R. felis từ chó được nhóm lại trong một nhánh với các trình tự tham chiếu của cùng một loài, được hỗ trợ bởi giá trị bootstrap 70% (Hình 6).

Các trình tự đại diện được sản xuất trong nghiên cứu này đã được lưu trữ trong cơ sở dữ liệu Genbank với các số đăng ký sau:
- Rhipicephalus sanguineus: PP389595 và PP389596
- Babesia vogeli: PP377903–PP377905
- Hepatozoon canis: PP373785
- Mycoplasma wenyonii: PP392702–PP392704
- Rickettsia felis: PP393500–PP393501
Kết luận
Nghiên cứu đã phát hiện rằng bọ chét chó nâu (R. sanguineus s.l.) là phổ biến ở chó tại các khu vực nghiên cứu ở Việt Nam. Nghiên cứu này đã xác thực nhiều tham số góp phần vào khả năng bị ký sinh trùng ở chó, điều này rất cần thiết cho các liệu pháp thú y thành công. Dữ liệu cho thấy sự hiện diện của R. felis, một tác nhân gây sốt phát ban, trong bọ chét R. sanguineus ở Việt Nam lần đầu tiên, điều này đặt ra những mối quan tâm lớn về sức khỏe cho động vật.
Từ khoá: chó, bọ chét, nguyên nhân gây bọ chét, yếu tố rủi ro, Việt Nam.
Link bài báo: https://www.frontiersin.org/journals/veterinary-science/articles/10.3389/fvets.2024.1435441/full#tab5

 English
English